
Dr Jalil Ahmad Bhatti
I have always found the virgin birth of Jesus, peace be on him, to be a fascinating topic. Basic human biology dictates that every healthy human’s genome is made up of 46 chromosomes in every cell: 22 identical pairs (44 chromosomes) and two gender-specific XX or XY chromosomes. A female body develops from the 46, XX and the male from the 46, XY genome. Complicating the matter of the virgin birth further; how did Mary, peace be upon her, a woman with a XX genotype, give birth to Jesusas with a XY genotype?
The claim that Maryas gave birth to Jesusas without a father is made principally by the religious texts of the Quran and the Bible. When searching for clues as to how this happened, our first assessment should be of the holy books. Interestingly, when we look at the events in the Quran in particular, our attention is drawn to the fact that not only was the birth of Jesusas described, but also Mary’s too:
“Remember when the woman of Imran said, ‘My Lord, I have vowed to Thee what is in my womb to be dedicated to Thy service. So do accept it of me; verily, Thou alone art All-Hearing, All-Knowing.’ But when she was delivered of it, she said, ‘My Lord, I am delivered of a female’ — and Allah knew best what she had brought forth and the male she was thinking of was not like the female she had brought forth — ‘and I have named her Mary, and I commit her and her offspring to Thy protection from Satan, the rejected.’” (Surah Aal-e-Imran, Ch3: V.36-37)1
“And remember her who preserved her chastity; so We breathed into her of Our word and We made her and her son a Sign for peoples.” (Surah al-Anbiya, Ch.21: V.92)
“And the example of Mary, the daughter of Imran, who guarded her private parts — so We breathed into him of Our Spirit — and she fulfilled in her person the words of her Lord and His Books and was one of the obedient.” (Surah al-Tahrim. Ch.66: V.13)
If we look at verses from Surah al-Anbiya and al-Tahrim, Maryas has been described with the gender pronouns of “her” and “him”, respectively. What significance does this hold? How could this possibly be related to the immaculate conception? Does it refer to Mary’s physiological or genetic uniqueness, by her possessing aspects of male and female elements? Historically, the pronoun “him” in this verse refers to Jesus; could it possibly allude to the nature of Mary’s biological gender instead? Interestingly, Jesusas is also called Isa ibn Maryam, meaning “Jesus, son of Mary” – an epithet usually related to the father, further indicating that she was both father and mother to Jesusas.
We should also remember that Maryas later gave birth to many of Jesus’ siblings. She was, therefore, a healthy woman, able to bear children following the immaculate conception of Jesusas.
As such, when looking at the mechanisms behind the virgin birth, we should bear in mind the following, when seeking its explanation:
- She must have been a virgin during the conception of Jesusas
- She was still capable of bearing children following the immaculate conception through the universal process of human reproduction
- Maryas must have had the ability to produce both male and female genetic material (i.e. 46, XY and 46, XX Chromosomes), despite being female in appearance
In the book Christianity – a Journey from Fact to Fiction, Hazrat Mirza Tahir Ahmadrh the Fourth Head of the Ahmadiyya Muslim Community detailed how the virgin birth could have occurred through parthenogenesis, hermaphroditism or as a combination of both. He explained as follows:
Hermaphroditism
“This term applies when organs of both sexes are present within a single female and the chromosomes show both male and female characters aligned side by side. Laboratory tests have revealed cases such as that of a hermaphrodite rabbit, which at one stage served several females and sired more than 250 young of both sexes, while at another stage became pregnant in isolation and gave birth to seven healthy young of both sexes.2 When autopsied, it showed two functional ovaries and two infertile testes, while in a pregnant condition. Recent studies suggest that such a phenomenon is possible, rarely, among humans also.”3
Parthenogenesis
“This is the asexual development of an ovum [egg] into an individual, without the aid of a male agent. It is observed among many lower forms of life such as aphids and also [some] fish. There is also evidence that parthenogenesis can be a successful strategy among lizards living under low and unpredictable rainfall conditions. In laboratory conditions, mice and rabbit embryos have been developed parthenogenetically to a stage equivalent to halfway through pregnancy, but have then been aborted. In recent study, human embryos could be activated occasionally by parthenogenesis using calcium ionophore as a catalyst. Such research raises the prospect that some early human pregnancy losses may have involved the parthenogenetic activation of the embryo.”3
But these terms – parthenogenesis and hermaphroditism – are general terms that encompass within them various potential mechanisms. This article aims to review plausible mechanisms in an unbiased, methodical manner.
Miracles and science
As explained by the founder of the Ahmadiyya Muslim Community, Hazrat Mirza Ghulam Ahmad, peace be on him, miracles such as the virgin birth do not denote a contravention to the law of nature, but the operation of an act of law of nature that we are unaware of:
“The truth of the matter is that God Almighty does not do anything against the laws of nature. What He does is that He creates the causes, whether we know of them or not, and these are always present. Hence, miracles, such as splitting of the moon and the fire [not burning Abraham, on whom be peace] are also no exceptions. Rather, they too resulted from some very subtle and hidden means and were based on true and factual science. Short-sighted people and those who are enamoured of dark philosophy cannot comprehend these things.”4
With scientific advances, potential answers as to what the “hidden means” were and what “factual science” underpins miracles, are illuminated. With the aid of modern developments in genetics and embryology, the virgin birth can be investigated and probed.
Before we do so, however, we need to understand a few definitions, particularly relating to the different mechanisms by which such an immaculate conception could have occurred.
Definitions
Genotype and phenotype
Genotype stands for the genetic makeup of the organism, enabling the development of a feature or trait. Phenotype is the expressed and observable trait or appearance caused by the genetic makeup of the organism.5
Germ cell
A germ cell can be an oocyte (ovum/egg) or a spermatozoa (sperm). The sperm comes from the male parent and the egg by the female parent. Each germ cell has typically half the amount of genetic material, meaning half the amount of chromosomes, and once joined, form a fully-functioning set of chromosomes.
Any human germ cell with 23 chromosomes is considered monoploid or haploid, i.e. it has a single set of unpaired chromosomes. Sperm can either have the genetic makeup of 23, X or 23, Y. The X and Y chromosome letters lead to sexual differentiation. An ovum has the default chromosomal make-up of 23, X. Once sperm and ovum fuse, the full set becomes 46, XX or 46, XY in a healthy female or male embryo respectively. This complete set of chromosomes is called diploid. A mismatch of chromosomes within the embryo, e.g. 45 or 47 instead 46 chromosomes are considered aneuploid. If a human embryo forms from multiple sets of 23 chromosomes, e.g. 69 (triploid) or 92 (tetraploid) chromosomes, it would be a specific case of aneuploidy called polyploidy.5–7
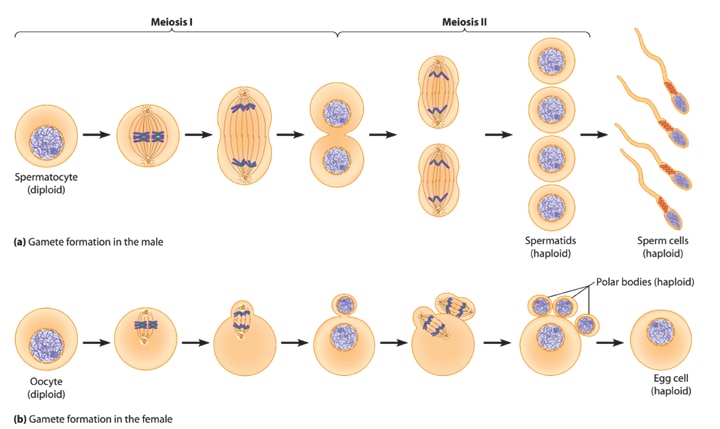
Meiosis
Meiosis is a part of the cell division cycle. Meiosis aims to create germ cells that are haploid, i.e. have half the amount of genetic material of the parent’s cells, so that when the sperm fertilises the ovum resulting in a full set of chromosomes. 7,9,10
Meiosis ensures variation in the genetic material in the germ cell compared to the parent cell. Causes for the differences in genetic makeup are independent re-assortment (much like shuffling a deck of cards) of the chromosomes, as well as homologous recombination (exchange of genetic material) between similar chromosomes.10 The result of normal meiosis leads to the creation of four germ cells in males and one ovum (with three polar/Barr bodies) in females.
A form of abnormal meiosis known as “non-disjunction”, is the failure of the parent cell to halve its genetic material and separate into daughter germ cells. The result may include a germ-cell with an extra or missing chromosome (aneuploidy); or multiple sets of all chromosomes in a single germ cell (euploid).9 Uniparental disomy is the condition when the child receives some or all of the genetic information from a single parent only. 11
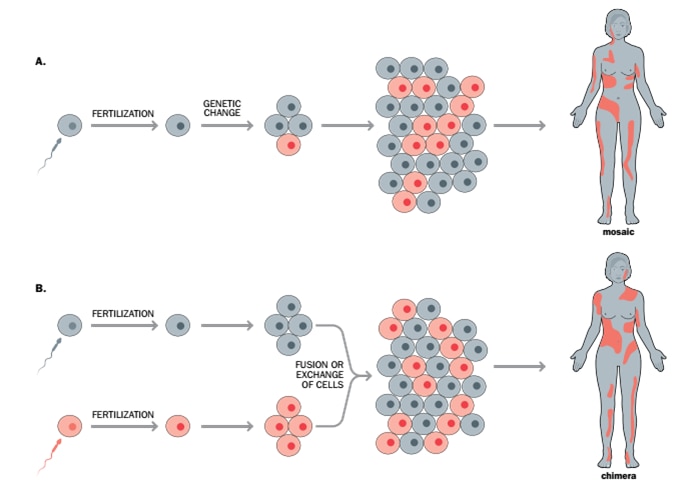
Mosaicism
Mosaicism is the presence of two or more populations of cell lines with different genotypes in a organism, originating from a single fertilised ovum. It can be a result of non-disjunction when during the process of cell division and proliferation from the original egg, non-disjunction of chromosomes leads to genetically similar but distinct cell lines in the eventual organism.
Another form of mosaicism can occur due to variation in genetic expression or even abnormality in chromosome numbers, e.g. cancerous cells, are derived from healthy cells, however their function changes due to distinct genetic changes. Therefore, the cancer cells in an individual have a different genotype compared to healthy cells due to somatic mosaicism, meaning that the individual has two genetically distinct but related cell lines from one original (progenitor) cell.
The relevance of mosaicism in this article is about the possibility of one progenitor cell changing to give rise to two different cell lines with genetic code for both male and female, enabling the development of gender-specific reproductive organs leading to the possibility of auto fertilisation.
The alternative role of mosaicism in the virgin mother is the formation of an egg with a distinct genetic code that has the parthenogenetic ability. The genetic code for the female phenotype in this egg has either been switched off or removed, and the remaining genetic code will give rise to a male child. 13
Germline mosaicism is the presence of different genetic make-up in germ cells compared to the genetic code of the rest of the organism, caused by abnormal development in the progenitor germ-cell line. The outcomes may include distinct chromosomal numbers of mosaic aneuploidy or mosaic euploidy. 5,7,9,11
Chimerism
A chimera is a single organism composed of cells from two or more fertilised eggs (known as zygotes). Occurring when sperms fertilise their congruent number of eggs (two or more) and join following fertilisation to form a single organism,14 thus two or more distinct cell lines of different genetic makeup create a single organism. In this case, fraternal twins would become “endogenous twins”, meaning that one twin would become incorporated into the second twin’s body, forming a sub-population of his or her cells. 7,9,10
X-linked gender (sex)-reversal
Patients that have X-linked Gender-reversal are genetically 46, XY (male) but present with a female phenotype appearance. Multiple genes orchestrate sexual differentiation, and under certain conditions, human females with 46, XY chromosomes were fertile and gave birth to children.16-19
Klinefelter’s syndrome
Klinefelter’s syndrome occurs in males typically when there are one or more additional X chromosomes, causing genotypes such as 47, XXY or 48, XXXY. Externally the person is male, however genetically the person has multiple numbers of the female X chromosome.5,7,20-23
On occasion, non-Klinefelter 47, XXY or 48,XXXY can occur, where a patient with multiple X chromosomes and a single Y chromosome will be female due to either abnormal X (single or multiple) or Y chromosome. 24-26
Epigenetics & epigenetic reprogramming
Epigenetics is the study of the mechanisms of external modification to the genetic code. So while the sequential nature of genetic code determines the intrinsic structure of genes, epigenetics looks at the expression of the code. Genetic code can be activated or deactivated by various mechanisms, resulting in distinct phenotypes from an unchanged original genetic code. Epigenetic reprogramming is a natural process but can also be induced artificially through scientific intervention. It has been utilised in stem cell research and animal cloning to change phenotype expression from an original master-copy genetic code, meaning the organism varies in appearances but is identical in genetic make-up.6
Applied epigenetic reprogramming
In an article published in Nature in 2002, embryonic cells derived from an adult female cat, and then transferred to a cat surrogate mother’s uterus. The surrogate mother underwent a healthy pregnancy and gave birth to the cloned kitten CC (short for either carbon-copy or copycat). This cloned kitten was distinctly different in appearance from her nuclear donor mother. Implying that even with identical genetic material, epigenetic reprogramming allows differentiation in phenotype and appearance.27
Having understood the different definitions, let’s look at the current theories on how the virgin birth could have taken place.
Current theories
Human chimera hypothesis
The current hypothesis of the virgin birth according to Dr Kemal Irmak, a professor in Histology and Embryology, relies on human chimerism 46,XY/46,XX as the main mechanism to produce a true hermaphrodite, who would then be capable of virgin birth.28,29
As we know, chimerism dictates the fertilisation of two eggs by two different sperm, thus creating a male and female foetus. The fusion of both fertilised eggs during the early phase of embryonic development, known as blastulation, would enable the female foetus to engulf the male foetus within itself, resulting in the creation of a single foetus with male and female cell lines. The developing embryo would have genotypes from both genders.14 The dominating genetic cell line expresses as the phenotype, in this case, female.
The individual cells 46, XX and 46, XY cells would correspond to their male and female reproductive organs on opposite sides of the abdomen.20,30 This mother would be a true hermaphrodite with internal ovaries, testes and possibly ovotestes.31 In all other aspects, she would be physiologically and anatomically female, allowing her to bear children through the normal process.
The virgin birth by a chimeric mother would occur by auto-fertilisation of the egg by a sperm, each provided by the separate reproductive organs within the same individual, producing a foetus in a virgin mother. Due to the presence of the Y chromosome from the internal testes, the resultant child could be a normal, healthy male.2
Foetus in Fetu Hypothesis
In the January 1927 Review of Religions article “The parthenogenesis” by Dr MS Nawaz, the hypothesis of “Foetus in fetu” was mentioned amongst other theories.32
Foetus in fetu can be considered a non-chimeric form of endogenous twins33,34, meaning that during the fetal development phase known as blastulation, the cells of one twin are absorbed by the other without fusing. If they fused, they would become chimeras. Instead, one foetus is subsumed into the other and arrests in the blastulation phase without serving any purpose to the host twin. The non-dominant subsumed twin is considered parasitic in nature and classified as a tumour usually.35 Parthenogenesis could happen if the host twin reaches puberty. The subsumed twin escapes the arrested (blastulation) phase, leading to the development of a foetus and eventual virgin birth.
New theories
XXY (non-Klinefelter) females and XY gender-reversal females
A Croatian woman with the male genotype 46, XY gave birth to a daughter with the same 46, XY genotype.17 Both mother and daughter received their Y chromosome from their respective fathers. The Y chromosome in the mother had a pathological variation, but in the case of the daughter, the Y chromosome was without any genetic defect. Regardless, both mother and daughter developed as females rather than males, as both had the condition of X-linked gender reversal. 24-26,36,37
Some of the family members of the mother had multiple disorders of sexual development. However, the mother had normal puberty, menarche, two spontaneous pregnancies and menopause. Conversely, the daughter was born infertile without a known cause despite the same 46, XY genotype as her mother.
The mother’s initial cell line during the blastulation stage diverged into four distinct cell lines. In the case of the daughter, the cell lines separated into three different cell types, indicating occurrence of non-disjunction genetic mosaicism in both the mother and the daughter.
The conception of the mother occurred by one sperm fertilising one egg. The genetic mosaicism enabled her to be fertile while expressing a majority of 46, XY chromosomes. Early non-disjunction during her conception caused her to be mosaic (but not chimeric), meaning that all her four cell lines originated from a single progenitor cell. The cell lines of her reproductive organs were 92.9% 46,XY and 0.6% 46,XX; the remainder made up of 45,X and 47,XXY.
The X-linked gender reversal condition in a female gives rise to three new potential scenarios of the Immaculate Conception of a male child:
Auto-fertilisation by Sertoli-Leydig cell tumours (arrhenoblastoma)
In specific individuals, a variation in the DICER1 gene can cause the formation of Sertoli-Leydig tumours from the ovarian cells, also known as arrhenoblastoma of the ovary.38 Sertoli and Leydig cells both are present in the male testis and are necessary for the formation of the sperm cell.7,39,40 Close analysis of these tumours has shown the potential of these tumours to create sperm cells in a 46, XX female.41–44 In this instance, a 46, XY female with Sertoli-Leydig tumour could undergo auto-fertilisation event via the arrhenoblastoma, releasing sperm to fertilise an egg from her ovaries, causing a virgin birth with a healthy male child.
Somatic Mosaicism with male and female reproductive systems (non-Klinefelter 47,XXY)
If, during the conception of the parthenogenetic mother, she had received an extra X chromosome from one parent, her genotype would be 47, XXY. Using the Croatian mother as an example, she had four distinct types of cell lines, namely 46,XX / 46,XY / 45,X and 47,XXY. 17,18,24-26
If the majority of her genotype was 47, XXY while being non-klinefelter, and during her development in the womb, a failure of the genetic material to separate by non-disjunction, the mother could have a minimum of two (or more) cell lines.
The Croatian mother was able to produce two following healthy diploid cell lines of 46, XX and 46, XY despite having a genetic make-up of 47, XXY.
If chimerism can cause the production of different reproductive organs on opposite sides of the abdomen, naturally mosaicism should be able to produce male and female reproductive organs on opposite sides of the abdomen as well.2
The 46, XX cell line will produce the female reproductive organs, including the ovary and oviducts, whereas the 46, XY cells will cause the development of testes. In this situation, the mother would be a true hermaphrodite with both male and female reproductive organs.
The fertilisation pathway would be the same as in the chimera, by auto-fertilisation. However, the main difference being that the conception of the mosaic mother involves one egg and one sperm, whereas in the chimeric mother, two sperms fertilising two eggs.2
Deactivation of X-linked gender reversal via germline mosaicism (uniparental disomy)
Another possibility, using the scenario of the Croatian woman to illustrate parthenogenesis, could be due to somatic mosaicism leading to germline mosaicism. To reiterate the mother had four distinct cell lines, she was fully fertile and expressed a female phenotype despite the presence of the Y chromosome. She had the condition called XY Gender (Sex)-Reversal.16,17,36,37,45
For the virgin birth to occur, one of her germ cells would require having a complete set of her genetic code with inactivation of the abnormal gene by various means. The Virgin Brith, in this instance, would be due to parthenogenesis and not autofertilisation.
This process would start in the 46, XY mother before the formation of the egg responsible for the virgin birth. During meiosis, recombination(a process of exchange and rearrangement of genetic material between closely related chromosomes) would lead to deletion or inactivation of the abnormal gene, the non-disjunction would result in an ovum with the full gamut (46,XY) of chromosomes and another ovum without any chromosomal information, this is called uniparental disomy.
The arrangement of the genetic material of the 46,XY ovum of the foetus compared to the mothers could give rise to four possible scenarios.
- Deactivation of the XY gender reversal gene by deletion
- The remaining genetic code rearranged wholly or partially.
- The remaining genetic code does not undergo rearrangement and stays identical and congruent to the mother’s genetic code.
- Deactivation of the XY gender reversal gene by epigenetic reprogramming
- The complete genetic code rearranged wholly or partially.
- The complete genetic code does not undergo rearrangement and stays identical and congruent to the mother’s genetic code.
Inactivation of the cause for the XY-Gender reversal takes place. Spontaneous parthenogenesis leads to the formation of an embryo and birth of a male child.
Explanation of theories
Earlier, we mentioned that the criteria for virgin birth would require the mother to give birth without the aid of a father.28,29 The Quranic verses may imply that Maryas, was not a typical biological female.
She was different in some way, possibly relating to her biological phenotype, by her possessing genetic material for both genders as well as reproductive organs. Though all the hypotheses discussed so far fulfil the criteria of the phenomenon of parthenogenesis. We still need to consider the implications of the different parthenogenetic possibilities, to illustrate the genetic relationship of the mother to herself and to the child while taking anatomical makeup of the mother into account.
- The first hypothesis we have presented states that virgin birth may have occurred due to chimerism. The product of two fertilised eggs (one fertilised egg being male and the other being female) fusing to produce a single individual with two different cell lines in the body.7 Such an individual would be a true hermaphrodite with internal testes (via the male cell line) and ovaries (via the female cell line) while retaining female appearance and anatomy in all other regards.
The genetic relationship of the mother to herself would be that of endogenous twin-siblings, and the foetus would have partial genetic similarity to the male and female cell lines. The egg would be produced from the ovaries via chimeric female cell line, whereas the sperm provided by the testis of the male cell line. As the mother would be a hermaphrodite, she will fulfil the criteria required for the virgin birth.
- The second hypothesis of foetus in fetu does not fulfil the criteria of the mother having internal organs from both genders. In contemporary scientific literature, in most cases of foetus in fetu, the enshrouded twin acts in a parasitic and cancerous way towards the host.33,34 The genetic relationship of the fetus to the mother would be that of siblings, rather than a mother-child relationship. However, it would still be an immaculate conception and considered a virgin birth.
- The third hypothesis mentioned was of arrhenoblastoma that leads to the auto-fertilisation of an egg. In this case, the mother has the same genetic material in both reproductive organs, the ovarian, as well as testicular cells. They are identical in nature, but their function is differentiated.41-44 The mother’s relationship with the child would be a recombinant clone, meaning their genetic code would be partially congruent in sequence. Both the mother and the child would have the 46, XY genes, similar in genotype, but different in phenotype, fulfilling the described criteria for the virgin birth.
- For non-Klinefelter mosaicism hypothesis, the mother would have been born due to the union of a single egg and sperm.24-26 Her mosaic genotype could give rise to multiple cell lines potentially including but not exclusively to 46,XX and 46XY. She would be a female hermaphrodite. The genetic code would express their respective reproductive organs, ovaries by 46,XY and testes by 46,XX.
- When reaching the age of maturation, the virgin birth would be due to autofertilisation of an egg by a sperm, with both being genetically from the mother’s intrinsic genetic code.
- The genetic relationship of the mother to herself would be that of semi-identical twins, and the foetus relationship to the mother would be of a recombinant clone, as the genetic material would have reshuffled during germ cell development, fulfilling the criteria for hermaphroditism through mosaicism.
- The final hypothesis of the XY female having an egg with a complete set of chromosomes causing virgin birth. Initially, by pulling the entire chromosomes set (46,XY, instead of 23,X)into a single egg. Subsequently, the cause for X-linked sex-reversal is deactivated.
In such an instance, parthenogenesis could occur without requiring fertilisation by a sperm. Different chemicals have been researched to induce artificial oocyte (egg) activation, including calcium ionophore, ionomycin, strontium and ethanol.27,46,47 It is possible that instead of an artificial, a spontaneous parthenogenetic inducer lead to the activation of an egg. Four different types of eggs with the complete set of genetic information from the mother are possible.
- The first possibility is an egg, which has undergone independent assortment and homologous recombination, creating an egg with the same genetic material as the mother, excluding the abnormal gene causing the XY-gender reversal by deletion, creating an incomplete recombinant clone.
- The second possibility is the creation of an egg that has not undergone independent assortment and where the genetic material is an exact copy of the genetic material of the mother, the XY-Gender reversal gene was inactivated by deletion, creating an incomplete identical clone.
- The third possibility is that the egg has inherited the complete code of the mother. Independent assortment and recombination lead to shuffling of the genetic code; epigenetic reprogramming leads to the deactivation of the abnormal XY-gender reversal gene and activation of the genes responsible for male reproductive organs. In this instance, the relationship of the mother to the child would be a complete recombinant clone at the genetic level.
- The fourth possibility is that the egg has inherited the complete code of the mother in identical and congruent sequence. Meaning both codes are an exact copy of each other. However, epigenetic reprogramming leads to activation of the male reproductive organs and deactivation of the gene for female reproductive organs. In this instance, the relationship of the mother to the child would be that of a complete identical clone.
In all four egg scenarios, this would be a parthenogenetic event and would fulfil the criteria for the virgin birth.17,36,37,45,48,49
Conclusion
Discussed were the following virgin birth hypotheses, in this article:
- Human chimaera
- Foetus in fetu
- Arrhenoblastoma auto-fertilisation
- Autofertilisation in XXY non-Klinefelter female
- Deactivation of XY-linked gender reversal by epigenetic reprogramming or deletion of the responsible genes.
We can say that there are a variety of possible mechanisms that could have enabled the virgin birth. The writings of the Promised Messiahas and our Holy book, the Quran are the guiding light and principle in allowing a better understanding of the topic of the virgin birth. The aim has been to present the scientific options without bias and as plainly as possible. The truth of the matter is known only to God.
“Allah knows what every female bears, and what wombs diminish and what they cause to grow. And with Him everything has a proper measure. He is the Knower of the unseen and the seen, the Incomparably Great, the Most High.” (Surah ar-Rad, Ch:13: V.9-10)
References
1. All Quranic references from the English translation by Hazrat Maulvi Sher Alira
2. Frankenhuis M, Smith-Buijs C, De Boer L, Kloosterboer J. A case of combined hermaphroditism and autofertilisation in a domestic rabbit. Vet Rec. 1990;126(24):598-599.
3. Ahmad T. Christianity: A Journey from Facts to Fiction. Tilford, Surrey: Islam International Publications; 2006.
4. Ahmad G. The Essence of Islam Vol 5: Tilford, Surrey: Islam International Publications; 2007.
5. Pritchard DJ, Korf BR. Medical Genetics at a Glance. 3rd ed. Chichester, West Sussex: John Wiley & Sons; 2013.
6. Cibelli JB, ed. Principles of Cloning. Second edition. Amsterdam: Elsevier/AP; 2014.
7. Sadler TW. Langman’s Medical Embryology. 13th edition. Philadelphia: Wolters Kluwer; 2015.
8. Ehamberg. Haploid vs Diploid. https://commons.wikimedia.org/wiki/File:Haploid,_diploid_,triploid_and_tetraploid.svg. Accessed April 3, 2020.
9. Carlson BM. Human Embryology and Developmental Biology. 5th ed. Philadelphia, PA: Elsevier/Saunders; 2014.
10. Brown TA. Introduction to Genetics: A Molecular Approach. New York: Garland Science; 2012.
11. Conlin LK, Thiel BD, Bonnemann CG, et al. Mechanisms of mosaicism, chimerism and uniparental disomy identified by single nucleotide polymorphism array analysis. Hum Mol Genet. 2010;19(7):1263-1275.
12. Hardin J, Bertoni G. Becker’s World of the Cell. Nineth Edition. Boston: Pearson; 2016.
13. Daughtry B, Mitalipov S. Concise Review: Parthenote Stem Cells for Regenerative Medicine: Genetic, Epigenetic, and Developmental Features. STEM CELLS Transl Med. 2014;3(3):290-298.
14. Boklage CE. Embryogenesis of chimeras, twins and anterior midline asymmetries. Hum Reprod. 2005;21(3):579-591. doi:10.1093/humrep/dei370
15. Read A, Strachan, Tom. Human Molecular Genetics. 5th ed. Garland Science; 2018.
16. MacLaughlin DT, Donahoe PK. Sex determination and differentiation. N Engl J Med. 2004;350(4):367–378.
17. Dumic M, Lin-Su K, Leibel NI, et al. Report of Fertility in a Woman with a Predominantly 46,XY Karyotype in a Family with Multiple Disorders of Sexual Development. J Clin Endocrinol Metab. 2008;93(1):182-189. doi:10.1210/jc.2007-2155
18. Kucheria K, Mohapatra I, Ammini A, Bhargava V, McElreavey K. Clinical and DNA studies on 46, XY females with gonadal dysgenesis. A report of six cases. – PubMed – NCBI. J Reprod Med. 1996;41(4):263-266.
19. Hines RS, Tho SPT, Zhang YY, et al. Paternal somatic and germ-line mosaicism for a sex-determining region on Y (SRY) missense mutation leading to recurrent 46,XY sex reversal. Fertil Steril. 1997;67(4):675-679.
20. Zhang M, Fan H-T, Zheng H-S, Zhang Q-S, Feng S-Q, Li R-W. Clinical characteristics of men with non-mosaic Klinefelter syndrome in northeastern China: implications for genetic counseling. Genet Mol Res. 2015;14(3):10897-10904. doi:10.4238/2015.September.9.27
21. Groth KA, Skakkebæk A, Høst C, Gravholt CH, Bojesen A. Klinefelter Syndrome—A Clinical Update. J Clin Endocrinol Metab. 2013;98(1):20-30. doi:10.1210/jc.2012-2382
22. Visootsak J, Aylstock M, Graham JM. Klinefelter Syndrome and Its Variants: An Update and Review for the Primary Pediatrician. Clin Pediatr (Phila). 2001;40(12):639-651. doi:10.1177/000992280104001201
23. Schoenhaus SA, Lentz SE, Saber P, Munro MG, Kivnick S. Pregnancy in a hermaphrodite with a male-predominant mosaic karyotype. Fertil Steril. 2008;90(5):2016.e7-2016.e10. doi:10.1016/j.fertnstert.2008.01.104
24. Acar H, Taskapu H, Muslumanoglu MH, Çapar M. A New Female Case with 47,XXY Karyotype and SRY. Andrology. 2016;05(01).
25. Röttger S, Schiebel K, Senger G, Ebner S, Schempp W, Scherer G. An SRY-negative 47, XXY mother and daughter. Cytogenet Genome Res. 2001;91(1-4):204–207.
26. Schmid M, Guttenbach M, Enders H, Terruhn V. A 47,XXY female with unusual genitalia | SpringerLink. Hum Genet. 1992;90:346-349.
27. Jose de Carli G, Campos Pereira T. On human parthenogenesis. Med Hypotheses. 2017;106:57-60. doi:10.1016/j.mehy.2017.07.008
28. Irmak MK. Self-fertilization in human: Having a male embryo without a father. Med Hypotheses. 2010;75(5):448-451. doi:10.1016/j.mehy.2010.04.021
29. Irmak M. Embryological basis of the virgin birth of Jesus. J Exp Integr Med. 2014;4(2):147. doi:10.5455/jeim.060113.hp.011
30. Talerman, A, Verp, MS, Senekjian, E, Gilewski, T, Vogelzang, N. True hermaphrodite with bilateral ovotestes, bilateral gonadoblastomas and dysgerminomas, 46,XX/46,XY karyotype, and a successful pregnancy. Cancer. Dec 15;66(12):2668-2672.
31. Yordam N, Alikasifoglu A, Kandemir N, Caglar M, Balci S. True hermaphroditism: clinical features, genetic variants and gonadal histology. J Pediatr Endocrinol Metab. 2001;14(4):421-427.
32. Khan SN. The Parthenogenesis. Rev Relig. January 1927:17-23.
33. Coolen J, Bradshaw B, Bhargava R, Lees G, Demianczuk N. Fetus-in-fetu: confirmation of prenatal diagnosis with MRI. Prenat Diagn. 2007;27(1):73-76. doi:10.1002/pd.1628
34. Hoeffel CC, Nguyen KQ, Phan HT, et al. Fetus in fetu: a case report and literature review. Pediatrics. 2000;105(6):1335–1344. doi:10.1542/peds.105.6.1335
35. Gupta S, Singhal P, Arya N. Fetus-in-fetu: A rare congenital anomaly. J Surg Tech Case Rep. 2010;2(2):77. doi:10.4103/2006-8808.73621
36. Sarafoglou K, Ostrer H. Familial sex reversal: a review. J Clin Endocrinol Metab. 2000;85(2):483–493.
37. Mazen IM, Kamel AK, Mohamed AM, et al. Unique Karyotype: mos 46,X,dic(X;Y)(p22.33;p11.32)/ 45,X/45,dic(X;Y)(p22.33;p11.32) in an Egyptian Patient with Ovotesticular Disorder of Sexual Development. Sex Dev. 2013;7(5):235-243. doi:10.1159/000351039
38. Slade I, Bacchelli C, Davies H, et al. DICER1 syndrome: clarifying the diagnosis, clinical features and management implications of a pleiotropic tumour predisposition syndrome. J Med Genet. 2011;48(4):273-278. doi:10.1136/jmg.2010.083790
39. Kyurkchiev S, Gandolfi F, Hayrabedyan S, et al. Stem Cells in the Reproductive System. Am J Reprod Immunol. 2012;67(6):445-462.
40. Pereira SL, Rodrigues AS, Sousa MI, Correia M, Perestrelo T, Ramalho-Santos J. From gametogenesis and stem cells to cancer: common metabolic themes. Hum Reprod Update. 2014;20(6):924-943. doi:10.1093/humupd/dmu034
41. Chou H-H, Lai Y-M, Lai C-H, Hsueh S, Soong Y-K. Sertoli-Leydig cell tumour in an infertile patient after stimulated ovulation. Hum Reprod. 1997;12(5):1021–1023.
42. Fagouri H, Moussaoui DR, Kouach J, et al. Complete Androgen Insensitivity Syndrome with a Sertoli-Leydig Cell Tumor. J Pediatr Adolesc Gynecol. 2014;27(5):e113-e115. doi:10.1016/j.jpag.2013.08.007
43. Gowri V, Koliyadan SV, Al Hamdani A, Al Kindy N. Successful term pregnancies after laparoscopic excision of poorly differentiated Sertoli-Leydig cell tumor of the ovary. J Gynecol Oncol. 2012;23(3):201. doi:10.3802/jgo.2012.23.3.201
44. Sachdeva P, Arora R, Dubey C, Sukhija A, Daga M, Kumar Singh D. Sertoli–Leydig cell tumor: A rare ovarian neoplasm. Case report and review of literature. Gynecol Endocrinol. 2008;24(4):230-234. doi:10.1080/09513590801953465
45. Mekkawy M, Kamel A, El-Ruby M, et al. Isodicentric Y chromosomes in Egyptian patients with disorders of sex development (DSD). Am J Med Genet A. 2012;158A(7):1594-1603. doi:10.1002/ajmg.a.35487
46. Bos-Mikich A, Bressan FF, Ruggeri RR, Watanabe Y, Meirelles FV. Parthenogenesis and Human Assisted Reproduction. Stem Cells Int. 2016;2016:1-8. doi:10.1155/2016/1970843
47. Kharche SD, Birade HS. Parthenogenesis and activation of mammalian oocytes for in vitro embryo production: A review. Adv Biosci Biotechnol. 2013;04(02):170-182. doi:10.4236/abb.2013.42025
48. Inoue H, Nomura M, Yanase T, et al. A Rare Case of 46, XX True Hermaphroditism with Hidden Mosaicism with Sex-determining Region Y Chromosome-bearing Cells in the Gonads. Intern Med. 1998;37(5):467-471.
49. Val P, Swain A. Mechanisms of Disease: normal and abnormal gonadal development and sex determination in mammals. Nat Clin Pract Urol. 2005;2(12):616-627. doi:10.1038/ncpuro0354